毛有东课题组在《自然》发表蛋白质机器动力学研究的重大突破
2018年11月12日,jxfjxf官网登录人工微结构和介观物理国家重点实验室、前沿交叉学院定量生物学中心毛有东课题组在《自然》(Nature)杂志在线发表了题为“Cryo-EM
structures and dynamics of substrate-engaged human 26S proteasome(底物结合的人源26S蛋白酶体的冷冻电镜结构和动力学)”的长论文(Article),通过冷冻电子显微镜和机器学习技术的结合,解析了人源蛋白酶体26S在降解底物过程中的七种中间态构象的高分辨(2.8~3.6埃)精细原子结构,最好局部分辨率达2.5埃。这些三维结构展现了惊人的的时空连续性(Spatiotemporal
continuity),生动呈现了原子水平的蛋白酶体和底物相互作用的动态过程,首次实现了对AAA-ATPase激酶六聚马达分子内ATP水解全周循环的完整过程的原子水平观测和三维建模,发现三种不同的ATP水解协同反应模式,用于调控蛋白酶体的复杂多样的功能。论文解决了一系列长期悬而未决的重要科学问题,包括:(1)蛋白酶体如何进行泛素识别和去泛素化;(2)底物是如何与ATPase马达结合起来的;(3)底物转运是如何启动的;(4)ATPase马达如何将化学能转化为机械能,进而实现底物解折叠的协同动力学机制。这是Nature上发表的系统性地、优于3.6埃分辨率水平实验研究超大复合蛋白质机器的动力学过程和原理的第一篇论文,标志着冷冻电镜的发展终于开始进入期待已久的全原子动力学分析的新时代,被审稿人誉为相关领域的“里程碑”。Nature编辑部和审稿人对该论文发表极其重视,从9月11日投稿、审稿到11月12日正式Online,仅用掉2个月时间。(Nature论文在线链接:https://www.nature.com/articles/s41586-018-0736-4)。
泛素-蛋白酶体体系(Ubiquitin-Proteasome
System,简称UPS)是细胞内最重要的蛋白质降解通路,对维持生物体内蛋白质的浓度平衡,以及对调控蛋白、错误折叠或受到损伤的蛋白的快速降解起着至关重要的作用,参与了细胞周期、基因表达调控等多种细胞进程,由UPS失常引发的蛋白质新陈代谢异常与众多人类重大疾病直接相关。2004年,Aaron
Ciechanover, Irwin Rose和Avram Hershko三位科学家被授予了诺贝尔化学奖,以表彰他们对该降解通路的发现。UPS中蛋白酶体是细胞中最基本的、最重要的不可或缺的、最为复杂的大型全酶超分子复合机器之一,人源蛋白酶体全酶包含至少33种不同的亚基,总原子质量约为2.5MDa。美国FDA批准的多种治疗癌症的药物分子即以蛋白酶体为直接靶标。近年来,随着冷冻电镜技术的发展和应用,人们对这一大分子机器的结构和功能研究得以不断深入。2016年,毛有东课题组与合作者报道了人源蛋白酶体基态的3.6Å冷冻电镜结构及其他三个亚纳米分辨构象,并首次发现一个亚稳态构象的核心颗粒(Core Particle,简称CP)底物转运通道处于开放状态(见PNAS
2016, 113: 12991-12996)。2018年4月,该课题组又报道了6个ATPγS结合状态下的26S动态结构,包括三个CP开放态对应的亚稳简并态近原子分辨(4~5Å)结构(见Nature Communications 2018, 9:
1360)。尽管这些工作揭示了蛋白酶体的基本架构和内在运动行为,但由于缺乏蛋白酶体与底物之间的相互作用,人们对于蛋白酶体如何实现底物降解的原子水平工作机制仍一无所知。此外,尽管冷冻电镜技术近年来广泛应用于分析具有动态特征的蛋白复合体结构和平衡态构象,但对其中间态结构和非平衡构象分析的分辨率水平往往局限在4~6埃或更低,离真正的全原子水平动力学分析还有相当一段距离。
为了真正实现原子水平的蛋白酶体底物降解动态过程的冷冻电镜三维重建和动力学表征,毛有东课题组攻克了两大技术难题。其一,如何在蛋白酶体完成底物降解之前抓到它的所有可能的中间态构象?课题组发展了一种新颖的核酸置换法,利用ATPγS降低AAA-ATPase激酶水解活性的特点,在底物降解中间过程,通过将ATP快速置换成ATPγS,结合快速冷冻的优势,从而扑捉到蛋白酶体在底物降解过程的中间态。其二,如何在从冷冻电镜数据中分析出更多构象的同时,还把分辨率做到3埃甚至更好?课题组通过多年持续努力,发展了多种基于人工智能和机器学习的冷冻电镜图像聚类的新型算法,并针对蛋白酶体的动力学特征,设计了一套极其有效的整合了多种算法的多构象分类流程。通过这两套技术方案的完美结合,课题组成功解析了人源蛋白酶体在降解底物过程中的七种不同的、但差别甚微的、高分辨原子水平的天然态构象(Native
states),完整展示了蛋白酶体从泛素结合到去泛素化,再到底物转运的动态过程。与同期在Science上发表的与底物结合的酵母蛋白酶体的4.2-4.7埃冷冻电镜结构(Science
doi:
10.1126/science.aav0725,来自加州伯克利分校和Scripps研究所)相比,该Nature论文不仅总构象数量多一倍,全部构象分辨率还高1-2埃。由于Science论文采用了抑制Rpn11去泛素活性的策略,其非天然态结构中底物并不能真正自由转运,所推测的机理仅限于底物转运这一步,对于其他三大Nature论文所回答重要问题均无法给出答案。这体现了该Nature论文不仅在实验方法的原创性上和数据分析水平和质量上,更在科学发现和问题探究的深度和广度上大幅超越了来自Science的竞争性论文。
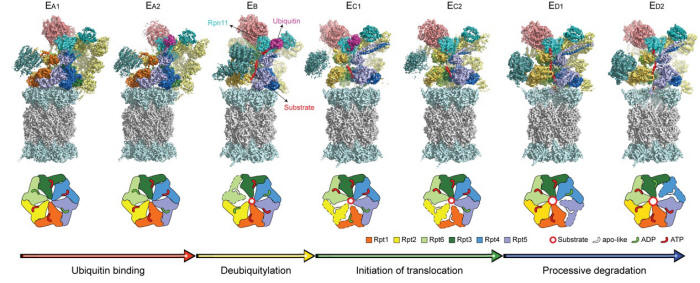
|
图一 七个利用冷冻电镜解析的精细原子结构完整揭示了从泛素识别、去泛素化反应、转运启动和持续降解的核心功能动态过程。
|
作为整个蛋白酶体的动力来源与运转核心,AAA-ATPase激酶分子马达展现出了三种不同的核苷酸水解协作模式,6个ATPase亚基协调工作,交替与底物发生相互作用。在去泛素化过程(EB态)中,处于对立位置的两个ATPase亚基Rpt2与Rpt4水解ATP,而Rpt5与Rpt6则释放ADP,ATPase内的底物转运通道被打开,使得底物可以进入轴心通道;与此同时,去泛素化酶Rpn11亚基与泛素及底物发生相互作用,执行其作为去泛素化酶的功能;在转运起始过程(EC态)中,相邻的两个ATPase亚基Rpt1与Rpt5会同时水解ATP,调控颗粒(Regulatory
Particle,简称RP)发生大规模转动并释放泛素;在底物去折叠与转运过程(ED态)中,三个相邻的ATPase亚基会分别同步进行ATP的结合、ADP的释放与ATP的水解,这一过程会单向传递下去,将ATP水解释放的化学能转换为机械能,使得相应的ATPase亚基发生刚体转动,推动底物的去折叠和单向输运,同时CP的转运通道入口打开,底物被送入通道中进行降解。这些研究结果为几十年来对蛋白酶体功能的研究提供了宝贵的第一手原子结构和动力学信息,对于理解生物体内蛋白质的降解过程和一系列负责物质输运的ATPase马达分子的一般工作原理具有极为重要的科学意义。
课题组博士后董原辰与博士生张书文为本论文的共同第一作者,毛有东为通讯作者。该论文的全部冷冻电镜数据在北大冷冻电镜平台上完成采集,大部分数据分析工作在北大高性能计算平台上完成。该论文的发表标准着北大冷冻电镜平台的建设后来居上,在数据采集效率和成像分辨率等各方面,已达到国际领先水平,具备了相当的国际竞争力。该研究项目得到了国家自然科学基金委、北大-清华生命科学联合中心、美国Intel公司并行计算研究基金、美国国家健康研究院、Edward
Mallinckrodt基金会的资助。


